What did G. H. Hardy, of Hardy-Weinberg fame, actually show?
Introduction
G. H. Hardy was a very successful mathematician of the early 20th century. Even today he is highly esteemed among professional mathematicians. He made seminal contributions to a variety of subdisciplines, including number theory and analysis, and he was among the most important British mathematicians of his day. His immortalization in the title of the founding theorem of population genetics therefore seems quite appropriate. But as is well-known among experts, it's actually quite ironic.
To start with, Hardy's contribution to the Hardy-Weinberg theorem was nothing more than a trivial mathematical observation expressed in a letter to the editor in Science. Two short paragraphs in the 1-page letter sufficed to convey the substance of the argument. The remainder is devoted to a contrived example and an expression of astonishment that "biologists" were not aware of something so straightforward. In fact, some biologists already knew of the result. In Germany, Wilhelm Weinberg [3] made the same observation at about the same time, but even he wasn't the first. Karl Pearson was. But Hardy was led to the belief that biologists were in the dark by an uncharacteristically misbegotten notion published by G. Udny Yule [1], who expressed a mistaken intuition, common among neophytes, that any dominant trait would eventually come to represent 3/4 of the population if the trait was not under selection. Hardy's motivation was to show that Yule was wrong, which required nothing more sophisticated than grade-school algebra. So the brilliant mathematician forever became linked by name to a mathematically trivial result.
And mathematically trivial it is. But one should not confuse mathematical simplicity with biological triviality. The Hardy-Weinberg theorem quite rightly serves as the foundation for population genetics, and among other things provides us a tool to identify traits that are evolving in real populations. So it is, I think, important to understand precisely what Hardy contributed to the theory.
Hardy's Result
In 1908, one approached problems in population genetics--the seeds of which had just been sown--often from different perspectives than the modern one. In this case, Hardy focused on absolute numbers of individuals of each genotype and neither on their frequencies (despite subsequent misinterpretations) nor the allele frequencies, as we tend to do now. So, first we'll look at his result using his notation and argument, and then demonstrate how it can be obtained more directly with modern ideas.
Hardy defined the numbers of individuals with genotypes AA, Aa and aa as p, 2q and r, respectively. Keep in mind that p and q do not mean what we typically teach in modern textbooks. These are numbers of individuals, not proportions. So, the total number of individuals in the population is p + 2q +r. If mating is random, the population is really large (or we ignore statistical fluctuations), sex ratios for each genotype are all unity, and there's no selection, then the number of matings among each genotype can be obtained by expanding
(1) (p + 2q + r)2,
and the number of individuals of each genotype in the next generation can be obtained by segregating their expected proportions. If we work it all out (which Hardy left the reader to do), we obtain the results in the following Table:
![Table 1: Mating frequencies as expressed by Hardy [2].](/sites/default/files/inline-images/Table-1.gif)
Now we just use Table 1 to partition up the mating frequencies to determine the number of each genotype in the next generation. For example, let pn be the number of AA individuals in generation n, and we want pn+1. We know that all offspring from AAxAA matings will be AA, while only half of the offspring from AAxAa matings and 1/4 of those from AaxAa will be AA. So,
pn+1 = pn2 + (1/2) (4pn qn) + (1/4) (4qn2) = (pn + qn)2.
This is the brute force approach. A more elegant way is to rewrite expression (1) as
(2) ((p+q) + (q+r))2.
If we square the first term, then we get the number of AA offpspring. Squaring the second term gives the number of aa offspring. The number of heterozygotes is the sum of the two cross-terms. Either way you look at it, you get the following
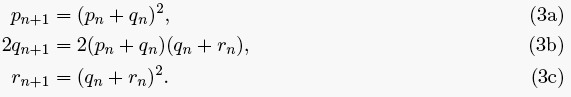
Hardy did none of this, nor did he quite use this notation. He assumed the reader grasped it without his help.
Our goal is to find a fixed point of system (3). Let that fixed point be (p, q, r). If Yule were right, then p + 2 q = 3r (there would be three times more dominant phenotypes than recessives at equilibrium). But, notice that a fixed point, by definition and equations (3a) and (3c), satisfies:

Substituting these results into equation (3b) tells us that at a fixed point we must also have
![]()
This evidently is not Yule's result. In addition, Hardy also stated without demonstration (because it was easy enough for the reader to figure out, or so he thought) that the numbers of each genotype would not change in successive generations following the first. Then he contrived an example to show in a concrete way how it worked.
The modern way of dealing with this is similar to Hardy's, although instead of numbers of each genotype, we focus on allele frequencies. So, beware of a change in notation here. Let p now represent the frequency of allele A and q be that of allele a. We assume these are the only alleles for the trait in the population, so p + q = 1. If mating is random, alleles are partitioned identically between the sexes--i.e., p is the same for both males and females--the population is large with no migration or selection, and mutation can be neglected, then the frequencies of all genotypes in the next generation are generated by the binomial square:
![]()
This is already a fixed point since, for any p,
![]()
and therefore qn+1 = qn. The genotype frequencies may shift in the first round of breeding, as Hardy noted, but the allele frequencies are fixed from the beginning. In the modern theory of dynamical systems, every point in the phase space, p in [0,1], is a neutrally stable fixed point. Where you start is where you stay forever.
One can also easily recover Hardy's condition, but before showing that let's disambiguate the notation. Let NAA, NAa and Naa be the number of individuals of each genotype. Hardy's observation was that "the condition for" system (3) to have a fixed point is

(Remember, in Hardy's notation, q is half the number of Aa individuals.) In the modern approach, random mating and all the other assumptions are expressed mathematically as the expansion of (p + q)2, so that the genotype frequencies after a single mating must be

![]()
In other words, random mating and all the other assumptions are true if and only if these are the genotype frequencies in all rounds of mating after the first. From here it is trivially easy to see that these frequencies hold if and only if
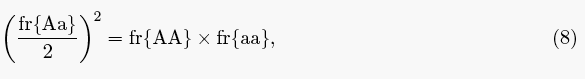
which is precisely Hardy's condition up to a constant equal to the population size. In summary then, if we are at an equilibrium for the genotypes, then condition (8) holds, and if condition (8) holds, we are at an equilibrium. Therefore, Hardy's somewhat cryptic, "it is easy to see that the condition for [the distribution being at an equilibrium] is q2 = pr" means that his condition is necessary and sufficient for an equilibrium.

Essay as a pdf is available here.